



La edad a la pubertad y primer parto afectan el desempeño reproductivo de la vaca de cría durante su vida productiva (Edwards et al., 2015; Lesmeister et al., 1973). Vaquillonas que se preñan en los primeros 21 días de servicio, permanecen durante más tiempo en el rodeo y destetan terneros más pesados respecto a las que se preñan más tarde (Perry y Cushman, 2013). La edad, peso vivo y condición corporal se asocian con una mayor probabilidad de alcanzar la pubertad y concebir más temprano (Engelken, 2008). Otros parámetros de crecimiento y desarrollo de las vaquillonas como la altura de anca y composición corporal, también han sido utilizados con éxito por su alta asociación con el peso vivo (Guggeri et al., 2014). Sin embargo, la pubertad es un proceso complejo que envuelve una serie de eventos. La nutrición, la edad y la genética influyen en el desencadenamiento de la pubertad, principalmente como reguladores de la maduración del eje hipotalámico – hipofisario – gonadal, la cual debe ocurrir para la iniciación de los ciclos estrales normales (Perry, 2016). La complejidad del inicio de la pubertad explica que la selección de vaquillonas de reemplazo basado únicamente en el peso vivo puede no ser suficiente. Más recientemente, se ha descrito al recuento máximo de folículos antrales (RMFA) y las concentraciones de hormona anti-Mülleriana (AMH) pre-servicio como marcadores fenotípicos de fertilidad en vaquillonas (Ireland et al., 2008). Las vaquillonas que tienen mayor número de folículos antrales tienen mayores niveles de AMH y progesterona y se preñan más temprano que vaquillonas con menor número de folículos antrales (Jimenez-Krassel et al., 2009; Martinez et al., 2016). En corderas se ha asociado un inicio más temprano de la pubertad con mayores concentraciones circulantes de AMH (Lahoz et al., 2012). La ocurrencia más temprana de la pubertad determina la sucesión de una mayor cantidad de ciclos estrales previo a la estación de cría, lo que se traduce en una mayor fertilidad al primer servicio (Byerley et al., 1987), y los mayores niveles de progesterona tienen un impacto positivo en el desarrollo embrionario (Diskin et al., 2006), contribuyendo a una concepción más temprana. A pesar de que el RMFA y la AMH son características fenotípicas que se expresan temprano y son repetibles en la vida del animal (Ireland et al., 2008), sus usos al destete como una herramienta para descartar vaquillonas menos fértiles no han sido extensamente evaluados (Morotti et al., 2017).
Las hipótesis de éste trabajo son: 1) el RMFA es repetible entre las evaluaciones al destete y pre-servicio, lo que permitiría seleccionar animales más fértiles a una edad más temprana; 2) vaquillonas con alto RMFA tienen parámetros de crecimiento y desarrollo compatibles con una pubertad más temprana, lo que puede estar relacionado con preñez más temprana; 3) el RMFA tiene una alta correlación con las concentraciones de AMH, por lo que ambos parámetros podrían ser utilizados de forma indistinta.
Los objetivos de éste trabajo fueron evaluar la repetibilidad del RMFA al destete y pre-servicio, su correlación con la AMH, parámetros de crecimiento y desarrollo, edad a la pubertad, concentraciones de progesterona en el ciclo pos-inseminación y preñez a los 13-15 meses en vaquillonas Hereford y Braford.
Materiales y métodos
Ubicación y animales
El protocolo experimental fue aprobado por el Comité de Ética para el Uso de Animales (CEUA, número de expediente 2013.13). Se utilizaron hembras Hereford y Braford en dos años distintos (2013-2014: Año 1 y 2015- 2016: Año 2), en un diseño experimental completamente al azar. El período de evaluación comenzó con la selección al destete de las terneras y finalizó con el diagnóstico de gestación, 30 días después del retiro de los toros del rodeo. El número de animales utilizados fue 42 Hereford y 40 Braford en el Año 1 y 50 animales de cada raza en el Año 2. Las terneras eran hijas de al menos 7 padres diferentes por raza. Las vaquillonas Hereford fueron manejadas siempre en la Unidad Experimental Glencoe y las vaquillonas Braford en la Unidad Experimental La Magnolia, ambas de INIA Tacuarembó.
Peso vivo, edad al inicio de las evaluaciones y alimentación
En el Año 1, los animales fueron seleccionados al destete en abril, con un peso vivo de 189 ± 2,2 kg y 189 ± 2,5 kg y 198 ± 3 días y 182 ± 3 días de edad, para hembras Hereford y Braford respectivamente. Desde el destete hasta mediados de setiembre las terneras pastorearon campo natural del ecosistema Campos (Allen et al., 2011) con una oferta de forraje de 10 kg de materia seca por kilo de peso vivo (kg MS / kg PV) (Sollenberger et al., 2005) y concentrado ad libitum en un comedero de autoconsumo. A partir de ahí, la dieta de las vaquillonas pasó a estar constituida por mejoramientos de campo natural durante 4 horas por día a una oferta forrajera de 7 kg MS / kg PV y el concentrado fue ajustado al 1% del peso vivo. El concentrado proporcionó 14 % de proteína bruta (PB) y 3 Mcal de energía metabolizable (EM) / kg MS. La ganancia promedio durante el período de recría fue de 0,700 ± 0,1 kg en las Hereford y de 0,681 ± 0,1 kg en las vaquillonas Braford, llegando al entore (a los 13-15 meses) con un peso promedio de 356 ± 3,5 kg y 322 ± 3,9 kg respectivamente. Desde el inicio hasta el final del período de servicio, las vaquillonas pastorearon campo natural con una asignación de forraje de 7 kg MS / kg de PV.
En el Año 2, las terneras fueron destetadas en marzo (Hereford) y abril (Braford), con un peso vivo de 164 ± 2,5 y 168 ± 3,6 kg de peso vivo y 165 ± 2 días y 174 ± 3 días de edad respectivamente. Durante la recría pastorearon verdeos de invierno (Raigrás y Avena) con una asignación de forraje promedio de 1,1 kg MS / kg de PV en ambos rodeos. Todas las hembras fueron suplementadas con granos secos de destilería con solubles (DDGS) de maíz (22,6 % de PB digestible y 2,8 Mcal/kg MS EM). Se comenzó con una suplementación a razón del 1,2 % del peso vivo para finalizar en 0,7 %. La ganancia promedio de la recría fue de 0,855 ± 0,1 kg en las Hereford y de 0,669 ± 0,1 kg en las Braford, llegando al entore (a los 13-15 meses) con un peso promedio de 378 ± 3,8 kg y 312 ± 4,6 kg en Hereford y Braford; respectivamente. Durante el servicio, los animales Hereford pastorearon campo natural y sorgo forrajero y los Braford Setaria, con una disponibilidad mínima de 7 kg MS /kg PV y 1 kg MS / kg PV, en pasturas naturales y anuales de verano; respectivamente.
Servicios
En el Año 1, todas las vaquillonas cíclicas se sincronizaron con el protocolo Ovsynch (Pursley et al., 1995) con el agregado de un dispositivo que contenía progesterona. Fueron inseminadas artificialmente a tiempo fijo (52 a 60 horas después de la administración de Cloprostenol), utilizando semen congelado de dos toros por raza con más del 60% de espermatozoides móviles después del descongelado. Diez días más tarde, se introdujo en cada rodeo un solo toro cuya aptitud reproductiva fue evaluada previamente, por un período adicional de 50 días.
En el Año 2, las vaquillonas fueron sincronizadas con prostaglandina F2? sintética (Cloprostenol-D, 150 mg i/m, Dalmaprost® Laboratorio Fatro, Uruguay), realizándose la inseminación a celo visto por 5 días. Se utilizó semen congelado de dos toros por cada raza, bajo las mismas condiciones que en el Año 1. Finalizada la inseminación, se introdujo en cada rodeo un solo toro cuya aptitud reproductiva fue evaluada previamente, por un período adicional de 50 días. Los inseminadores fueron dos técnicos entrenados y trabajaron en ambos rodeos y años.
Determinaciones en los animales
Recuento de folículos antrales
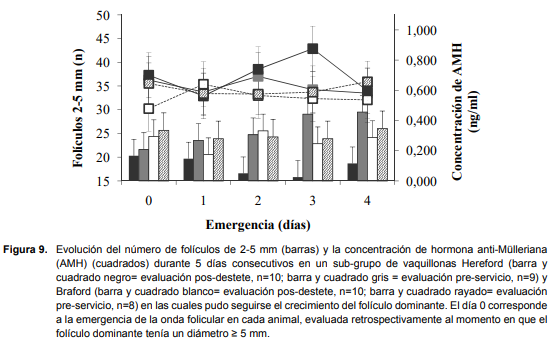
Se evaluó el recuento de folículos antrales (RFA) en ambos ovarios, registrándose en mapas ováricos el diámetro, número y posición de todos los folículos ? 2 mm. Las evaluaciones se llevaron a cabo en dos momentos (pos-destete = 246 días de edad promedio y pre-servicio = 385 días de edad promedio) utilizando un ecógrafo de modo B en tiempo real y una sonda transrectal de 7,5 MHz (Aloka Co., Ltd., Tokio, Japón). El mismo operador realizó todas las mediciones en el mismo año, pero fueron diferentes entre años. En el Año 1 la evaluación se realizó día por medio, completando 6 días de evaluación para cada animal y para cada momento (pos-destete y pre-servicio), mientras que en el Año 2, se realizó durante 5 días consecutivos en ambos momentos. En el año 2, la ecografía diaria permitió definir la emergencia de una onda folicular en un total de 37 vaquillonas. La emergencia de la onda fue definida realizando el seguimiento retrospectivo del folículo dominante de esa onda, cuando tenía un diámetro ? 5 mm (Día 0= emergencia del folículo dominante). El recuento máximo de folículos antrales (RMFA) para cada animal, fue el día durante cada seguimiento (pos-destete y pre-servicio) en que se registró la mayor población total (suma de ambos ovarios) de folículos antrales ? 2 mm.
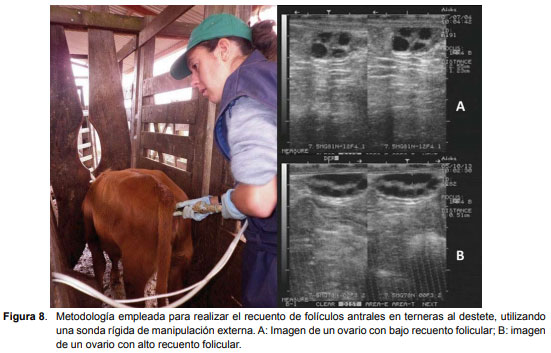
En la evaluación pos-destete se utilizó una sonda rígida que fue manipulada externamente (Figura 8) utilizando la metodología descrita por Viñoles et al. (2010), en la cual los ovarios son visualizados utilizando la vejiga y el útero como estructuras de referencia y rotando la sonda en el sentido de las agujas del reloj para localizar el ovario izquierdo y en sentido contrario a las agujas del reloj para localizar el derecho.
Para el seguimiento pre-servicio los animales fueron sincronizados con dos dosis de prostaglandina F2? sintética (Cloprostenol-D, 150 mg i/m, Dalmaprost® Laboratorio Fatro, Uruguay) administradas con un intervalo de 11 días. La evaluación comenzó 48 horas después de la segunda dosis y en este caso se utilizó una sonda flexible, palpándose los ovarios y colocándolos directamente debajo de la sonda.
Muestreos sanguíneos y determinaciones hormonales
Se recolectaron muestras sanguíneas por venopunción yugular todos los días en los momentos de evaluación de la cuenta folicular, y entre ambos momentos la frecuencia de muestreos sanguíneos se redujo a dos veces por semana. La recolección de sangre se interrumpió en aquellas vaquillonas que habían alcanzado la pubertad evidenciado por la presencia de un cuerpo lúteo en la evaluación pre-servicio, pero en las restantes se continuó hasta el final del período de servicio. Este patrón de muestreos fue definido para: 1) determinar la concentración de AMH el día del RMFA en ambos momentos; y 2) conocer con la mayor exactitud posible la fecha de pubertad de cada vaquillona. El suero fue separado por centrifugación durante 10 minutos a 3500 rpm y las muestras se almacenaron a -20°C hasta la determinación de progesterona y AMH.
Las determinaciones hormonales se realizaron en el Laboratorio de Endocrinología y Metabolismo Animal, Facultad de Veterinaria, Montevideo, Uruguay. Las concentraciones de progesterona fueron determinadas por un radioinmunoensayo (RIA) en fase sólida. En el Año 1 se utilizaron kits DPC (Diagnóstico Product Co., Los Ángeles, CA, EEUU). La sensibilidad del ensayo fue de 0,079 ng/ mL y el coeficiente de variación intra-ensayo para los controles de calidad bajo (1 ng/mL) y alto (29 ng/mL) fueron 10% y 8%; respectivamente. En el Año 2 se utilizaron kits de MP (BIOMEDICALS, INC. Solon, OH 44139 USA), la concentración mínima detectable del ensayo fue de 0,1 ng/ml y los coeficientes de variación intra-ensayo para el control bajo (0,94 ng/ml) y el control alto (5,09 ng/ml) fueron 13,6 % y 13,4%; respectivamente.
La concentración de AMH fue determinada solamente el día del RMFA para cada animal en cada evaluación y en los cinco días de evaluación en aquellos animales que se pudo observar la emergencia de la onda folicular (n = 37). Se utilizó el kit de ELISA AL-114 (Ansh Labs, Texas, EEUU), el cual es diseñado para el uso específico en vacas y fue previamente validado en vaquillonas Braford (Bo et al., 2016) y Hereford (Sequeira et al., 2014). La sensibilidad del ensayo fue de 0,011 ng/mL. En el Año 1 el coeficiente de variación intra-ensayo para los controles de calidad bajo (0,177 ng/mL) y alto (0,379 ng/ mL) fueron de 15,7% y 6,8%; respectivamente. Mientras que en el Año 2, los coeficientes de variación intra-ensayo para el control bajo (0,259 ng/ml) y el control alto (0,946 ng/ml) fueron 20,7% y 17,0%; respectivamente.
Parámetros reproductivos
El inicio de la pubertad fue determinado usando las mediciones de progesterona. Una sola muestra con concentraciones ? 1 ng/ml o dos muestras consecutivas con concentraciones ? 0,5 ng/ml fueron indicativas de actividad luteal y por tanto de la pubertad.
Se realizó el diagnóstico de gestación y la determinación de la edad embrionaria/fetal cada 30 días, desde el inicio del servicio hasta 30 días después de retirados los toros. Se utilizó un ecógrafo Agroscan con un transductor transrectal lineal dual de 5,0/7,5 MHz (Biotay SA, Montevideo, Uruguay). A su vez, se determinaron la fecha de concepción, la edad a la concepción y el momento de concepción. La fecha predicha de concepción se calculó restando la edad del embrión o feto a la fecha del diagnóstico de gestación. Utilizando la fecha de concepción y la fecha de nacimiento de cada vaquillona, se calculó su edad en el momento de la concepción. El momento de la concepción fue definido como los días desde el inicio del servicio en que la vaquillona quedó preñada.
Parámetros de crecimiento y desarrollo
Peso vivo
El peso vivo se determinó en todos los animales cada 14 días, desde el inicio del experimento (destete) hasta el final del entore. La medición se realizó siempre por la mañana y utilizando la misma balanza (True test GR 3000s, True-test Corporation Limited, Montevideo, Uruguay). Usando esta información se calculó la ganancia diaria de peso vivo.
Altura de anca
La altura de anca se midió con una regla justo encima de los huesos de la cadera, con los animales parados sobre una superficie plana, la semana siguiente al seguimiento folicular pos-destete y pre-servicio.
Composición corporal
Se estimó mediante ecografía el espesor de grasa (grasa subcutánea que recubre el músculo dorsal largo entre la 12° y 13° costilla, medida en mm), área de ojo de bife (AOB, área del músculo dorsal largo medido en cm2 ) y el espesor de la grasa en la grupa (espesor de grasa subcutánea en el sitio P8 expresado en mm) el día de medición de la altura de anca. La ecografía de composición corporal fue realizada utilizando un escáner modo B en tiempo real, con un transductor lineal de 3,5 MHz (Aloka Co., Ltd., Tokio, Japón) y se almacenaron en una PC portátil (Houghton y Turlington, 1992). Las imágenes capturadas en la PC fueron analizadas utilizando el software Biosoft® (Biotronics Inc., Ames, Iowa, EEUU).
Análisis estadísticos
El impacto del RMFA en explicar las variables de interés (crecimiento y desarrollo, edad a la pubertad) se estudió utilizando el dato individual en cada evaluación, para cada raza y año. Esta definición se basó en la alta variabilidad existente en el RMFA entre vaquillonas y la gran variación en los criterios para la clasificación en alto, medio y bajo RMFA (Morotti et al., 2015). En éste trabajo se clasificó a los animales en bajo, medio y alto RMFA utilizando puntos de corte que dividieron a cada población de animales, en cada evaluación y año, en tres tercios. Esta clasificación solamente se utilizó para describir variables relacionadas a la fertilidad, cuando el análisis utilizando el dato de RMFA individual estuvo relacionado con las variables de interés.
Todos los análisis fueron realizados utilizando el Statistical Analysis Software (SAS 9.4, SAS Institute Inc, Cary, Carolina del Norte, EEUU, 2002). Las variables continuas y discretas fueron analizadas mediante modelos lineales generalizados y generalizados mixtos, respectivamente, utilizando los efectos fijos RMFA o clase de RMFA, observación y la interacción entre estos factores. El análisis de los residuales de las variables RMFA y AMH mostró el rechazo de la hipótesis de normalidad (procedimiento univariado, test de Shapiro-Wilk, P < 0,05), por lo cual se realizó la transformación logarítmica de los datos para ambas variables. La repetibilidad del RMFA y las concentraciones de AMH medidas con un intervalo de 6 meses (evaluaciones pos-destete y pre-servicio) se estudió mediante análisis de varianza usando la fórmula r = S2 A / (S2 + S2 A), donde S2 A es la varianza entre grupos y S2es la varianza dentro del grupo. Adicionalmente, se estudió la correlación entre el RMFA y las concentraciones de AMH en ambas evaluaciones, para cada raza y año. Únicamente en el Año 2 en que el seguimiento ecográfico se realizó en forma diaria, se determinó la consistencia de las medidas del recuento de folículos antrales (RFA) (folículos de 2 a 5 mm) y de las concentraciones de AMH para cada evaluación (pos-destete y pre-servicio) y raza (Hereford y Braford) en un subgrupo de vaquillonas (n = 37) en los que se detectó la emergencia de una onda folicular. Para evaluar la relación del RMFA y la concentración de AMH con la edad a la pubertad, y del RMFA con peso vivo, altura y el AOB, se realizaron regresiones múltiples creándose una línea de tendencia y una ecuación de regresión. Las gráficas de regresión se realizaron usando JMP 10 (SAS Institute Inc., Cary, NC, USA).
La clasificación en RMFA bajo (? 21 folículos), medio (22 – 31 folículos) y alto (? 32 folículos) solamente fue utilizada para las vaquillonas Braford, en la evaluación posdestete en el año 2 para evaluar las concentraciones de progesterona en el ciclo posinseminación y la frecuencia de animales inseminados y preñados a la inseminación artificial y la preñez final. Todos los modelos fueron considerados significativos si P < 0,05, y las tendencias fueron identificadas cuando 0,05 < P ? 0,10.
Resultados
Repetibilidad del RMFA pos-destete y pre-servicio
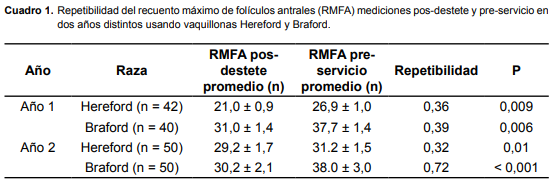
En el cuadro 1 se presenta la repetibilidad del RMFA para la cuenta pos-destete y preservicio por año y por raza. La repetibilidad más alta se observó en el Año 2, para las vaquillonas Braford.
Repetibilidad de la concentración de AMH
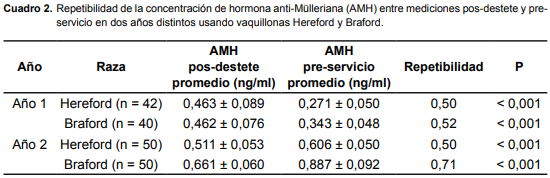
En el cuadro 2 se presenta la repetibilidad de la concentración de AMH entre las mediciones pos-destete y pre-servicio. La repetibilidad para la concentración de AMH tuvo un comportamiento similar al RMFA, siendo mayor en el Año 2, en las Braford.
Correlación del RMFA y la concentración de AMH
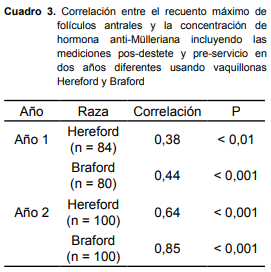
La correlación entre el RMFA y la concentración de AMH para cada año y raza, incluyendo los valores pos-destete y pre-servicio de cada animal, se presenta en el cuadro 3. En todos los casos la relación entre ambas variables fue significativa (P < 0,001), pero las correlaciones fueron mayores para las vaquillonas Braford en el Año 2.
Consistencia de las mediciones del RFA y AMH durante evaluaciones diarias
En el Año 2, en que las mediciones de RFA y AMH fueron realizadas durante 5 días consecutivos, se evaluó la consistencia de las mismas para ambos momentos (pos-destete y pre-servicio) y para ambas razas (Hereford y Braford) utilizando un subgrupo de vaquillonas (n = 37). La consistencia del recuento de folículos de 2 – 5 mm en la evaluación pos-destete fue 0,87 (n = 10) para las vaquillonas Hereford y 0,94 (n = 10) para las Braford y en la evaluación pre-servicio de 0,79 (n = 9) y 0,93 (n = 8) para las vaquillonas Hereford y Braford, respectivamente.
La consistencia de las mediciones de AMH en cada evaluación (pos-destete y pre-servicio) fue evaluada utilizando el mismo subgrupo de vaquillonas. En Hereford, la consistencia para la evaluación pos-destete y pre-servicio fue 0,89 (n = 10) y 0,95 (n = 9) respectivamente; y en Braford para la evaluación pos-destete fue 0,92 (n = 10) y para la pre-servicio 0,94 (n = 8). Se observó mínima variación diaria en las concentraciones de AMH de este subgrupo (P = 0,45; Figura 9), y mínima variación diaria en el número de folículos de 2 – 5 mm (P = 0,4; Figura 9).
Relación RMFA con parámetros reproductivos
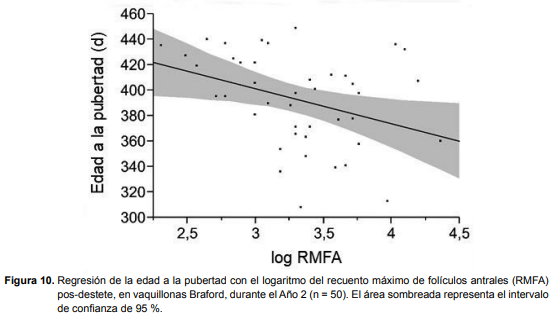
En el Año 1, en las vaquillonas Hereford y Braford, y en el Año 2 en las Hereford, las evaluaciones pos-destete no evidenciaron relación entre el RMFA y parámetros reproductivos (edad a la pubertad, momento de concepción, edad a la concepción y preñez final). Sin embargo, en el Año 2, en las vaquillonas Braford la evaluación pos-destete evidenció que animales con mayor RMFA alcanzaron la pubertad a una edad más temprana (r2 = 0,129; P = 0,0196; Figura 10). La ecuación que mejor describe ésta relación es pubertad = 483,79704 – 27,503385 * log RMFA. La edad promedio a la pubertad de cada grupo (bajo = 418,7 ± 9,0 días; medio = 379,2 ± 8,4 días y alto = 383,4 ± 8,7 días) determinó que una menor proporción de vaquillonas de bajo RMFA fueran inseminadas al inicio del período de servicios (2/16), compradas con las de RMFA medio (12/17) y alto (7/17; P < 0,01). Sin embargo, no se observaron diferencias en la proporción de vaquillonas preñadas a la inseminación, el momento, ni la edad a la concepción entre grupos (bajo RMFA = 0/2, 28,6 ± 4,9 días y 429,9 ± 7,1 días; medio RMFA = 6/11, 23,0 ± 4,4 días y 419,1 ± 6,5 días; alto RMFA= 4/7, 24,3 ± 4,2 días y 431,2 ± 6,5 días; respectivamente; P > 0,05). La preñez final fue similar para las vaquillonas de bajo (11/16), medio (13/17) y alto (13/17) RMFA (P > 0,05).
En la evaluación pre-servicio no se encontraron asociaciones entre RMFA y variables reproductivas en ninguno de los años ni razas.
Relación entre la concentración de AMH y los parámetros reproductivos
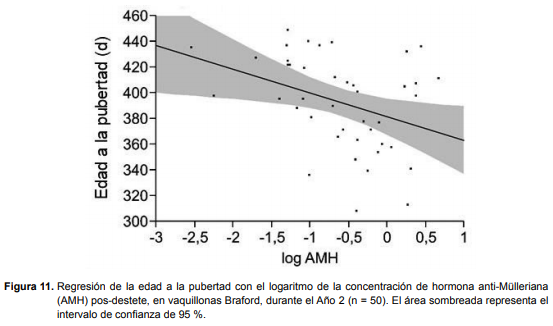
En las vaquillonas Braford durante el Año 2, se observó una relación entre el logaritmo de AMH y la edad a la pubertad, tal que a mayores concentraciones de AMH, menor edad a la pubertad (r2 = 0,135; P = 0,0168; Figura 11). La ecuación que mejor describe ésta relación es pubertad= 381,56734 – 18,446481 * log AMH.
Relación del RMFA con las concentraciones de progesterona
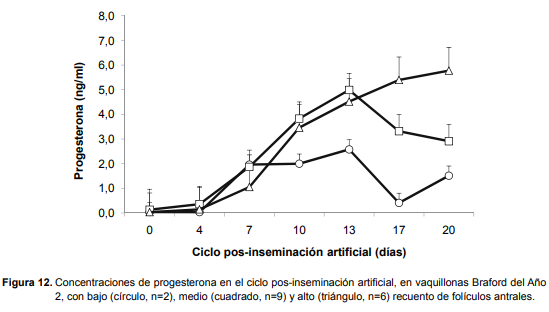
En las vaquillonas Braford del Año 2, las concentraciones de progesterona en el ciclo pos-inseminación estuvieron afectadas por la clase de RMFA, el día y la interacción de ambas (P < 0,001), dado que las terneras de clase baja tuvieron menores concentraciones de progesterona a partir del día 10 del ciclo comparado con aquellas de clase media y alta (Figura 12).
Relación RMFA con parámetros de crecimiento y desarrollo
En el Año 1, no se encontraron asociaciones entre RMFA y los parámetros de desarrollo y composición corporal, en ninguna de las evaluaciones, para ninguna de las razas. Sin embargo, en el Año 2, las asociaciones entre RMFA y los parámetros de desarrollo y composición corporal se observaron en las vaquillonas Braford, en la evaluación posdestete. La disminución del RMFA se correspondió con menor peso (r2= 0,146; P < 0,01), altura (r2 = 0,159; P < 0,01) y AOB (r2 = 0,111; P = 0,018; Figura 13). Las ecuaciones que describen éstas relaciones son peso = 122,9377 + 23,796473 * log RMFA, altura anca = 97,320422 + 3,6574946 * log RMFA y AOB = 26,445724 + 4,2351617 * log RMFA.
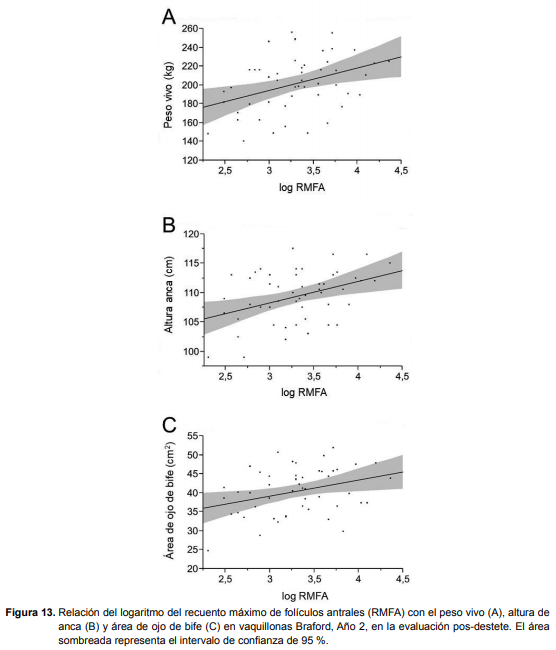
En el mismo año y con la misma raza pero en la evaluación pre-servicio, las vaquillonas con menos RMFA siguieron siendo más bajas (P = 0,0042) y tuvieron menos AOB (P = 0,0307).
Discusión
Este es uno de los primeros trabajos que usa el RMFA al destete como herramienta de selección precoz de reemplazos. Las hipótesis testeadas en este estudio fueron parcialmente aceptadas. El RMFA fue repetible entre las evaluaciones pos-destete y pre-servicio solamente en uno de cuatro casos. En ese único caso (vaquillonas Braford del Año 2) en que la repetibilidad fue alta, encontramos una relación entre el RMFA y parámetros que describen el crecimiento y desarrollo, por lo que las vaquillonas con menor RMFA crecieron de forma más lenta y alcanzaron la pubertad más tarde. Además, en las vaquillonas Braford del año 2 se encontró una alta correlación entre el RMFA y la concentración de AMH, sugiriendo que ambas variables pueden utilizarse de manera indistinta para seleccionar hembras al destete. Nuestros resultados refuerzan el concepto de que el inicio de la pubertad es un proceso complejo que se ve afectado entre otros factores por el peso vivo, la tasa de crecimiento y la raza; y que por esto una única medida del RMFA o de la concentración de AMH al destete pueden no ser técnicas útiles para seleccionar vaquillonas más fértiles, por lo menos en la raza Hereford.
La repetibilidad del RMFA observada en vaquillonas Braford en el Año 2, fue muy buena aunque no tan alta como la previamente reportada por Burns et al., (2005); Rico et al., (2009) y Souza et al., (2015) (0,87- 0,95). Este resultado positivo indicaría que para este caso, la selección de hembras al destete puede ser posible, sin embargo, nuestros resultados fueron pobres en los restantes casos (Año 1, vaquillonas Hereford y Braford y Año 2, vaquillonas Hereford). Los pobres resultados podrían estar relacionados con la metodología utilizada en los diferentes experimentos y al momento fisiológico en que se encontraban los animales (pre vs peri-púberes). En la evaluación pos-destete se utilizó una sonda rígida, de manipulación externa, mientras que en la evaluación pre-servicio se utilizó una sonda flexible, y esto podría estar afectando la repetibilidad obtenida en este estudio. Los ovarios, en la evaluación pos-destete, fueron visualizados utilizando la vejiga y el útero como estructuras de referencia y rotando la sonda en sentido horario para ubicar al ovario izquierdo y antihorario para ubicar el ovario derecho, de la misma forma que lo descrito para la oveja (Viñoles et al., 2010). Sin embargo, en la evaluación pre-servicio los ovarios se palparon, y se colocaron debajo de la sonda, obteniéndose una mayor definición de su estructura anatómica. La menor repetibilidad del RMFA obtenida utilizando la sonda rígida en animales peri-púberes ha sido reportada previamente (Lents et al., 2014). Otro factor que puede estar afectando los resultados es que la repetibilidad fue medida considerando solamente el día de máxima cuenta folicular (RMFA), en dos mediciones separadas por 6 meses, en animales pre-púberes en la primer medición, y peri-púberes en la segunda. Los resultados de repetibilidad presentados por Ireland et al., (2008) son de animales púberes sincronizados, en que se evaluó la cuenta máxima en ondas sucesivas del ciclo estral, realizando dos ecografías diarias. Esta diferencia metodológica es relevante por dos motivos: 1) los complejos cambios hormonales que ocurren alrededor de la pubertad, pueden estar afectando secundariamente la transición y la dinámica del pool de folículos antrales (Yelich et al., 1996); 2) la probabilidad de que dos mediciones sean iguales es mayor cuando son más cercanas en el tiempo (Littell et al., 1998). Sin embargo, investigaciones recientes (Morotti et al., 2017) reportaron en ganado Braford alta repetibilidades (0,90 a 0,92) del RMFA entre mediciones al destete y al año de edad. Esto refuerza los buenos resultados obtenidos en las vaquillonas Braford del Año 2, e indica que la realización del RMFA al destete puede ser una herramienta que permita predecir la performance reproductiva en esta raza al servicio.
Un resultado relevante de éste trabajo tiene que ver con el momento del ciclo en el que se debe realizar la evaluación del RFA. En nuestro experimento, las vaquillonas fueron examinadas ultrasonográficamente por 5 a 10 días, con el objetivo de capturar la emergencia de al menos una onda folicular. Las primeras publicaciones en esta área fueron realizadas conociendo el día de la emergencia de la onda (Burns et al., 2005), pero la metodología fue modificada para simplificar su uso en condiciones de campo (Martinez et al., 2016). Gobikrushanth et al. (2017) observaron una baja repetibilidad (0,37) entre la cuenta folicular realizada en un único día conociendo o no el momento del ciclo. Sin embargo, nuestros resultados indican que un única medición en un momento desconocido del ciclo estral sería posible y permitiría una mayor aplicación productiva. En el Año 2, el número de folículos de 2 – 5 mm evaluados durante 5 días consecutivos, fue altamente consistente, particularmente en las hembras Braford. La mínima variación diaria del número de folículos de esta clase está en concordancia con la descripción de las ondas foliculares en vacas (Rajakoski, 1960) y ovejas (Bartlewski et al., 1999), lo que refuerza el concepto de que una única medida en un día desconocido del ciclo estral es también correcta desde el punto de vista biológico.
Solamente en vaquillonas Braford en el segundo año de evaluación, el RMFA pos-destete permitió identificar las vaquillonas que alcanzarían la pubertad antes. Este resultado puede estar explicado por dos factores: 1) diferencias raciales en edad a la pubertad, ya que Bos Indicus y sus cruzas alcanzan la pubertad más tarde (Nogueira, 2004) , y 2) diferencias en los planos nutricionales entre años y razas, lo cual provocó menores tasas de ganancia diaria y peso vivo pre-servicio en las vaquillonas Braford. Ambos, nutrición y genética se combinaron para estimular una pubertad más temprana en las vaquillonas Hereford, las cuales estaban más cerca del límite biológico para esta raza. Debido a que más vaquillonas Hereford al momento de la evaluación pre-servicio estaban en el período peri-puberal, los cambios hormonales y su impacto en el crecimiento folicular durante este período fisiológico, podrían estar impidiendo lograr una buena repetibilidad entre ambas mediciones (pos-destete y pre-servicio). Sin embargo, en las vaquillonas Braford existe margen de mejora en el plano nutricional por lo que el uso de una herramienta adicional como el RMFA puede haber colaborado en la selección de hembras más precoces. En el Año 2, vaquillonas Braford con menos RMFA tuvieron menor crecimiento y desarrollo, pudiendo esto explicar la demora en alcanzar la pubertad y la menor proporción de vaquillonas inseminadas. A su vez, esas vaquillonas con reducido número de folículos antrales, tuvieron menos concentraciones de progesterona en el ciclo posinseminación. Esto no resultó, sin embargo, en diferencias en el momento de concepción o en el porcentaje de preñez final. Trabajos previos en ganado para carne no encontraron una relación beneficiosa entre el RMFA y la pubertad (Cushman et al., 2014; Mossa et al., 2013), pero si se ha descrito que vaquillonas con alto RMFA tienen mayores concentraciones de progesterona (presente estudio, (Jimenez-Krassel et al., 2009)). Además, vaquillonas con mayor RFA presentan un mayor tamaño uterino y contenido de proteína uterina, lo que indica la existencia de un ambiente más auspicioso para el desarrollo embrionario, variables que podrían explicar la concepción más temprana respecto a vaquillonas con menor RFA (Cushman et al., 2014; Jimenez-Krassel et al., 2009; Martinez et al., 2016; McNeel et al., 2017; McNeel y Cushman, 2015). El incremento en las concentraciones de progesterona en vaquillonas con alto RMFA después del día 10 del ciclo estral, podría contribuir a un incremento en el elongamiento del embrión y favorecer el reconocimiento materno de la preñez (Diskin y Niswender, 1989). Es importante considerar que estos son los primeros datos generados en nuestras condiciones, y con un número muy reducido de vaquillonas, particularmente en la clase de bajo RMFA. En el mismo sentido, la evaluación de variables reproductivas como el momento de concepción y la tasa de preñez final requieren de un número elevado de vaquillonas, lo que podría explicar en parte las similitudes entre las distintas clases de RMFA en este estudio. Por lo tanto, se requiere más estudios para confirmar la asociación entre bajas concentraciones de progesterona en vaquillonas Braford con bajo RMFA y su relación con la fertilidad.
La relación entre RMFA y parámetros que describen el crecimiento y desarrollo fue también observada solamente en las vaquillonas Braford en el Año 2. Los resultados sugieren que vaquillonas con menor RMFA fueron más livianas, tuvieron menor AOB y fueron más bajas, que las de medio y alto RMFA. Estos resultados concuerdan con datos publicados anteriormente (Eborn et al., 2013), y sugieren que la nutrición pre y pos-natal podrían afectar la reserva ovárica (Amundson et al., 2015; Evans et al., 2012; Mossa et al., 2013). La asociación de los parámetros de crecimiento y desarrollo con la edad a la pubertad (Patterson et al., 1992) sugiere que vaquillonas con un mayor potencial de crecimiento alcanzan el porcentaje de peso adulto crítico para comenzar a ciclar a una edad más temprana. Sin embargo, se debe tener la precaución de evitar la selección de animales excesivamente grandes y pesados, que podría tener consecuencias negativas en la fertilidad, particularmente considerando las restricciones anuales en el consumo de energía en sistemas pastoriles (Do Carmo et al., 2016; Jenkins, 2009). Por este motivo no se recomienda el uso únicamente del peso vivo como herramienta para seleccionar vaquillonas de reemplazo, sino que es importante considerar la fecha de nacimiento de esos animales (Roberts et al., 2017).
En nuestro experimento, el RMFA y la AMH se correlacionaron positivamente en todos los análisis, aunque con coeficientes menores a los reportados previamente (Ireland et al., 2008). La correlación observada en el año 2 en ambas razas fue buena, pero no en el año 1. Además, la repetibilidad de la concentración de AMH promedio fue mayor a la observada con el RMFA, siendo moderada en tres casos (Año 1, Hereford y Braford y Año 2, Hereford) y muy buena en el restante, Braford del Año 2 (0,71). Estos resultados coinciden con reportes previos (Rico et al., 2009) e indican que la AMH podría ser usada para seleccionar al destete las vaquillonas Braford más precoces. Inicialmente, se planteó que una de las ventajas más importantes del uso de la AMH respecto al RMFA, era su estabilidad a lo largo del ciclo estral (Ireland et al., 2008). Sin embargo, trabajos posteriores y el nuestro demuestran que el RFA (contando folículos ? 2 mm), puede realizarse en cualquier momento del ciclo estral. Los folículos de 2 mm, representan un alto porcentaje de los folículos antrales presentes en el ovario (Rajakoski, 1960), y no serían afectados por las variaciones en la concentración de FSH como los folículos ? 3 mm (Martinez et al., 2016). Otros aspectos relevantes a favor del uso del RFA son: la información se obtiene al pie del animal, permite evaluar el desarrollo del tracto reproductivo en vaquillonas pre-servicio y su costo es menor que el del análisis de AMH (3,5 vs 9,7 dólares por animal sin tener en cuenta los honorarios del laboratorio). Sumado a esto, se han reportado importantes variaciones en la concentración de AMH para la misma muestra en el mismo laboratorio (Cushman, com. pers; Sanderson, com. pers), por lo que se requieren ajustes en la metodología del análisis de AMH. La asociación entre ambas variables sugiere que podrían ser utilizadas de manera indistinta para seleccionar al destete aquellas vaquillonas Braford que consecuentemente alcanzarían la pubertad tempranamente, aunque sería útil tener en cuenta las ventajas y desventajas de cada alternativa, como fue previamente desarrollado.
Conclusión
Los resultados de estos experimentos muestran que el RMFA y la concentración de AMH no son útiles para la selección de hembras Hereford pos-destete o pre-servicio. Sin embargo, el RMFA y la concentración de AMH pos-destete podrían ser utilizados para seleccionar vaquillonas Braford que alcanzan la pubertad a una menor edad. Adicionalmente, la evaluación puede ser realizada en cualquier día del ciclo estral, haciendo esta técnica aplicable a los sistemas de producción de carne. Vaquillonas Braford que tienen un mayor RMFA tienen parámetros de crecimiento y desarrollo más acelerados, lo que puede contribuir con una pubertad más precoz. Estos resultados son relevantes para el sector criador, ya que permitiría eliminar del rodeo a vaquillonas menos fértiles a temprana edad (destete) que serán destinadas a la venta o al engorde con la consecuente mejora en el retorno económico de ésta actividad en varias fases del sistema productivo (ej. Rodeo de cría y feedlot). Sin embargo, se requieren más estudios con mayor número de vaquillonas Braford para probar la validez del RMFA y la AMH como herramientas de selección temprana. Estas técnicas no deben aplicarse en forma aislada, sino asociadas a un plano nutricional que estimule un adecuado crecimiento y desarrollo de las vaquillonas de reemplazo.




{kind=link}