



Una proporción sustancial de la producción de carne en Colombia se origina en sus extensas llanuras orientales. Sin embargo, en este escenario y en un contexto global, la demanda de producción ganadera requiere cada vez más que satisfaga las expectativas sociales y ambientales, además de ser económicamente eficiente. Un conjunto de datos que contiene registros de cinco años de sistemas de producción de vacas y terneros recolectados en el Centro de Investigación Carimagua ubicado en el Departamento de Meta fue interrogado retrospectivamente para comprender la dinámica de matriz de flujo derivada del peso vivo (LW) de las emisiones de metano (CH4). Las emisiones totales estimadas de CH4 (kg) durante el período de gestación fueron similares entre las vacas destetadas convencionales (CW; 37.86 ± 0.506 kg) y las destetadas temprano (EW; 37.47 ± 0.476 kg). Sin embargo, con un promedio de dos lactancias, las emisiones totales de CH4 fueron mayores (p <0.0001) en vacas CW (38.67 ± 0.456 kg) que en sus contrapartes EW (14.40 ± 0.435 kg). Las emisiones totales de gases desde el nacimiento hasta la edad comercial comparable fueron mayores (p <0.0001) para terneros CW (43.11 ± 0.498 kg) que para terneros EW (40.27 ± 0.472 kg). Se concluyó que los conjuntos de datos pastorales a mediano y largo plazo y las nuevas inquietudes son adecuados para comprender diferentes contextos y adaptaciones a las condiciones climáticas contemporáneas. Sin embargo, los sistemas agrícolas convencionales serán menos vulnerables al medio ambiente si las prácticas de manejo de EW involucran el uso estratégico y temporal de pasturas mejoradas. Los roles de la medicina veterinaria y las ciencias animales se discuten brevemente en el contexto de una variabilidad climática sin precedentes para proporcionar una guía para el futuro incierto.
Palabras clave: rebaños de carne, huella de carbono, peso vivo, emisiones de metano, destete.
INTRODUCCIÓN
Las llanuras orientales de Colombia incluyen 17 millones de hectáreas de sabanas neotropicales bien drenadas, 3.5 millones de las cuales tienen pendientes bajas y se pueden cultivar, mientras que 6.4 millones son montañosas, diseccionando paisajes que descansan sobre suelos de suelos variables, pero generalmente superficiales, excepto en pequeños valles que ocasionalmente se siembran con pasturas introducidas (Sánchez y Cochrane 1985). Aunque se entienden de manera imperfecta (Rippstein et al. 2001; Parr et al. 2014), estas extensas sabanas de pastizales (es decir, Llanos) son un elemento fuerte en la cultura colombiana, el discurso histórico y socioeconómico, a la vez que tienen un importante valor patrimonial, ecológico y geológico. para nativos, coloniales y otros habitantes multiétnicos (Navas Ríos 1999; Pulido et al. 2007).
Sin embargo, a pesar de la continua intensificación de las industrias agrícolas, culturales, pesqueras, forestales, mineras, recreativas y de ecoturismo (Castiblanco et al. 2015), la industria de la carne en sabanas neotropicales y pasturas sembradas ha capturado históricamente el uso principal de la tierra (Van Ausdal 2009; Rausch 2013; Huertas-Ramírez y Huertas-Herrera 2015). Una imagen similar, pero a una escala más amplia, es evidente dentro de los subsistemas biofísicos, socioeconómicos, humanos (pueblos indígenas y no indígenas) y culturales de los sistemas remotos de cría de carne en los frágiles pastizales del norte de Australia [pastizales, matorrales, bosques nativos y sabanas de bosques tropicales (Tothill y Gillies 1992; Marshall y Smajgl 2013; Marshall et al. 2014; Russell-Smith y Sangha 2018)].
En este contexto, y para satisfacer las aspiraciones de los medios de vida agrícolas y no agrícolas y mantener las estructuras sociales y comunitarias, es deseable preservar la identidad de los extensos rebaños de carne de cría. Esto requiere lidiar no solo con las eficiencias nutricionales y reproductivas, sino también adoptar las mejores prácticas con respecto al crecimiento y comercialización de destetados, calcetines y añores (Kleinhesterkamp y Habich 1985), lo que a su vez impulsa una amplia variación de rendimiento de un año a otro. salidas del sistema pastoral (Mejía et al. 2004).
Paralelamente, la producción sostenible y / o la intensificación sostenible de la industria de carne de res dependiente de los recursos sensibles en las sabanas constituye un desafío importante, dadas las interdependencias e interrelaciones entre, pero no limitadas a (i) el aumento de la transformación de la tierra (Smith et al. 2010; Gobierno australiano 2015); (ii) vulnerabilidad a la incertidumbre climática (Marshall et al. 2014; Ramos-Montaño y García-Conde 2016); resiliencia social y capacidad de adaptación de los productores primarios y las comunidades rurales al cambio climático (Marshall 2010; Marshall y Smajgl 2013); y la implementación requerida de un marco pastoral de mitigación extenso e integrado (Ramírez-Restrepo y Charmley 2015) para ayudar a la seguridad nutricional y los rendimientos financieros agrícolas positivos en el contexto de una creciente demanda mundial de calidad alimentaria [(CONPES) 2014; Thompson y Martin 2014; Norton 2017].
Como se sugirió anteriormente, llegar a mercados globales valiosos requiere cada vez más productos de calidad respaldados por una impresión documentada de bajo contenido de carbono (McAulifee et al.2018). Esto es consistente con Godde et al. (2019) y Tedeschi et al. (2002, 2019), quienes argumentaron que frente a la variabilidad climática es fundamental utilizar enfoques de modelado para capturar representaciones ganaderas ambientales cortas y largas, simples y complejas de sistemas agrícolas de la vida real anteriores o actuales en los pastizales tropicales de Australia o en otros lugares. Por lo tanto, los datos de referencia y los escenarios para diversas cargas ambientales de los sistemas de producción basados ??en carne de sabana en Colombia son deseables para integrar el intercambio profundo de conocimientos locales y el modelado basado en datos. Curiosamente, se sabe más sobre algunos impactos ecológicos de los pastos sembrados (Fisher et al.1994; Ramírez-Restrepo y Vera 2019) en este extenso ambiente de sabana neotropical, que de las sabanas nativas pastadas per se (Ramírez-Restrepo et al.2019).
El objetivo de este estudio fue cuantificar las emisiones de metano de carne de vacuno y ternera (CH4) y los índices de intensidad y eficiencia derivados durante dos ciclos reproductivos (CR) consecutivos en relación con el destete y la edad de un año, utilizando una matriz matemática de flujo de peso vivo individual ( LW) datos de un experimento de cinco años en el Centro de Investigación Carimagua (CRC) ubicado en los Llanos de Colombia.
MATERIALES Y MÉTODOS
Detalles de las condiciones del sitio
El Centro de Investigación Carimagua, ubicado en el Departamento de Meta, se encuentra en un Oxisoles de suelo franco arenoso o franco arcilloso con buen drenaje, con densidad aparente global, pH del suelo, fósforo disponible, materia orgánica del suelo y valores de saturación de aluminio de 1.4 g / cm3, 4.74, 2.48 mg / kg, 3,07% y 80%, respectivamente (Rao 1998; Fisher et al. 1994; Rao et al. 2001). La precipitación pluvial media y la temperatura ambiente durante el período de diciembre a marzo (27,4º C y 169 mm / m2) y la estación húmeda (26,2º C y 2,790 mm / m2) durante el período 1984-1987 se registraron estudios en el sitio meteorológico CRC ( 4º 36’44.6 ”de latitud N, 74º 08’42.2” de longitud oeste; Vera y Ramírez-Restrepo 2017).
Estudios utilizados para modelar
Los datos de peso vivo provienen de proyectos diseñados por R. R. Vera Infanzón e implementados por los dos autores en el contexto del antiguo Programa de Pastos Tropicales del Centro Internacional de Agricultura Tropical (CIAT) y financiados por su presupuesto central. Cubre dos RC completos y consecutivos y se repitió dos veces en años consecutivos provenientes de ensayos de pastoreo de sabana neotropical realizados éticamente por médicos de medicina veterinaria registrados en Colombia (Vera y Ramírez-Restrepo 2017). Los experimentos incluyeron parejas de vacas y terneros Brahman (Bos indicus) y cruzados Brahman x San Martinero (nativo; Bos taurus) sujetos a destete convencional (CW; 304 ± 2.29 días) y destete temprano (SE 114 ± 2.20 días).
La investigación original también sirvió para un propósito práctico de identificar en los rebaños EW, el valor alimenticio del pasto Andropogon gayanus en asociación con las leguminosas forrajeras Pueraria phaseoloides y Centrosema acutifolium durante 4.79 meses (30 días) y 4.85 meses hasta que las terneras CW fueron destetadas en la sabana en 1984 (Replicar 1) y 1985 (Replicar 2), respectivamente. Lo que también fue de interés fue conocer el impacto de la sabana en la producción cuando los reproductores (terneros destetados; 10.1 ± 1.71 meses) fueron monitoreados durante 14.46 meses adicionales para convertirse en un año (24.0 ± 0.05 meses). Esta combinación particular de ganado de cría pura y cruzada, forrajes, condiciones ambientales y de manejo retrata los actuales regímenes locales de extracción y agricultura extensiva más ampliamente reportados en otros lugares (Vera y Hoyos 2018).
Enfoque de modelado
Las fluctuaciones reproductivas, productivas, LW y los valores de SR estaban sujetos a una conceptualización mental y una representación matemática adjunta (Tedeschi 2019) de emisiones de CH4 y pérdidas de gas de energía (MJ) calculadas en un modelo mecanicista de hoja de cálculo Excel® como se describe en detalle por Ramírez-Restrepo y Vera (2019). Brevemente, las medidas relevantes incluyeron la ingesta ad libitum derivada de LW (2.1% del total de LW; Fisher et al. 1987) (DMI; Eq. 1) durante la concepción, gestación, parto, destete y primer período seco. Paralelamente, el sistema de emisión de CH4 derivado de LW después de 56 días de edad (Huws et al.2018) identificó el impacto de asignación de las emisiones diarias (g) (Ec. 2) asociadas con los indicadores clave de desempeño ambiental en términos de parámetros reproductivos, y índices de intensidad y equivalentes de dióxido de carbono (CO2) (CO2-eq) de eficiencia. Las emisiones fecales de CH4 y óxido nitroso (N2O) del estiércol y la orina del ganado, así como de CH4 y N2O del suelo están fuera de los cálculos actuales (Ramírez-Restrepo et al.2019).
Estas estimaciones ambientales utilizaron el potencial de calentamiento global del horizonte de 100 años de 34 para CH4 (Mueller y Mueller 2017), mientras que los datos de los toros no están incluidos en la matriz de flujo.
Ecuaciones de regresión predictiva:
Eq. 1. Ingesta de materia seca (Ramírez-Restrepo y Vera 2019).
Y = 0.014 (± 0.0036) (LW) + 2.216 (± 1.3156) r 2 = 0.491, p <0.01; CV = 18,94; r.s.d = 1.34; r = 0,70, p <0,01
Eq. 2. Metano g / d (Ramírez-Restrepo y Vera 2019).
Y = 0.324 (± 0.0577) (LW) + 16.176 (± 21.0879) r 2 = 0.663, p <0.0001; CV = 16,78; r.s.d = 30.82; r = 0,81, p <0,0001.
En caso de necesidades futuras, impulsadas por cambios extremos, los parámetros sensibles en el modelo pueden modificarse para simular rebaños sobre la base de la nueva estructura. Sin embargo, es esencial asegurar datos de entrada relevantes de condiciones tropicales extensas para lograr indicadores clave confiables de rendimiento ambiental y productivo.
análisis estadístico
La varianza asociada con las medias de tratamiento se analizó en SAS (2016) utilizando el procedimiento GLIMMIX para mediciones de LW, DMI y todas las emisiones de CH4 derivadas. Efectos independientes de las réplicas (RP, es decir, 1 y 2; a partir de los años 1984 y 1985, respectivamente), práctica de destete (es decir, CW y EW), RC (es decir, 1 y 2), las interacciones entre la práctica de destete y RC; y entre RP, la práctica de destete y RC se ajustaron en un modelo lineal usando el efecto aleatorio de la vaca.
El análisis se complementó con el procedimiento MIXED para evaluar los efectos lineales ajustados de la RP, la práctica de destete y la RP mediante la interacción de la práctica de destete en los períodos de concepción (seco) posteriores al destete. Los resultados se presentan como medias de mínimos cuadrados ± errores estándar de las medias (LSM ± SEM), a menos que se indique lo contrario. Se calcularon diferencias significativas al nivel de probabilidad de 0.05 y se muestran valores p específicos cuando están disponibles.
RESULTADOS
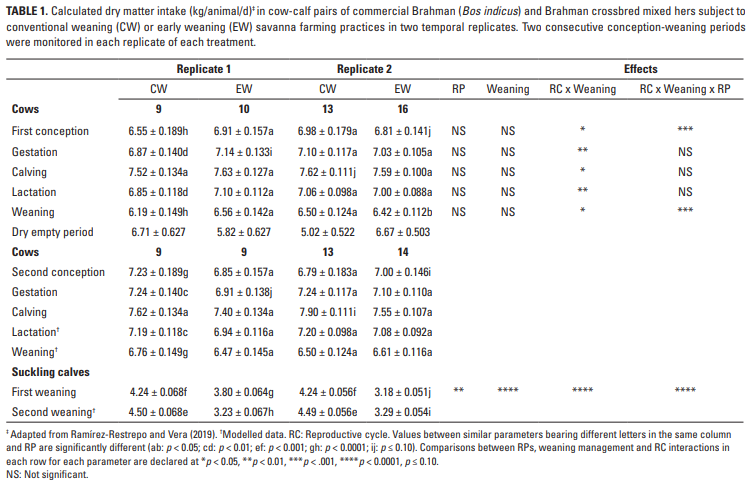
En las vacas, el DMI (kg / animal / d) se vio afectado por la interacción (p <0.05) entre RC y las prácticas de destete (Tabla 1). Sin embargo, la interacción entre la práctica de destete RC x RP y RP fue dominante (p <0.001) en la primera concepción y durante el período de vacío seco (Tabla 1). En general, durante los dos RC, el DMI en los tratamientos CW y EW fue similar durante la concepción (6.89 ± 0.114 vs 6.90 ± 0.107), gestación (7.11 ± 0.081 vs 7.04 ± 0.077), parto (7.66 ± 0.066 vs 7.54 ± 0.063), lactancia (7.08 ± 0.069 vs 7.03 ± 0.065) y destete (6.49 ± 0.090 vs 6.52 ± 0.084). El pastoreo común en las sabanas condujo a una variación en el DMI de los terneros entre RP (p <0.01) debido a los tratamientos de destete y en todas las interacciones (p <0.0001). En promedio, el DMI fue mayor (p <0.0001) en terneros CW (4.37 ± 0.037) en comparación con los terneros EW (3.38 ± 0.035).
En la concepción, las diferencias promedio de las emisiones de CH4 entre las vacas CW (123.05 ± 2.628 g / d) y EW (123.29 ± 2.462 g / d) no fueron significativamente diferentes, mientras que las emisiones generales fueron más bajas (p <0.05) en el primer RC (121.38 ± 1.919 g / d) que en el segundo RC (124.97 ± 1.943 g / d). Esto dio como resultado una interacción consistente (p <0.001) RC x destete x replicar interacción.
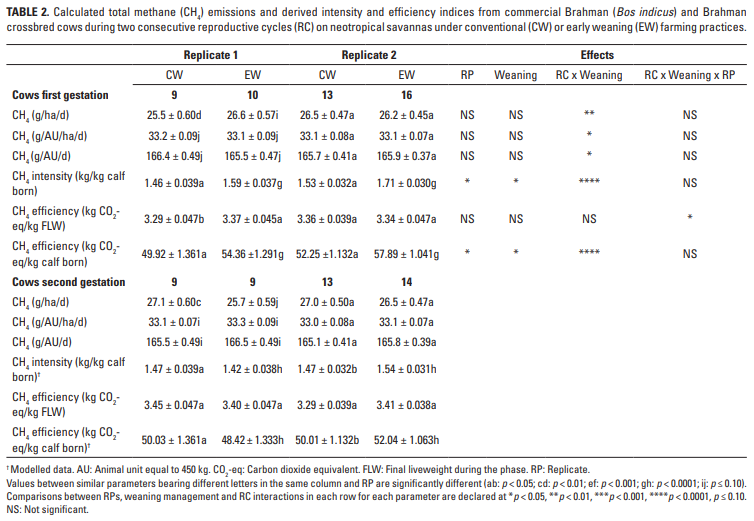
La Tabla 2 muestra las emisiones de CH4 de las vacas en términos de área, día, unidades animales e índices de intensidad y eficiencia durante las gestaciones de dos RC consecutivos. Las diferencias en las emisiones entre las RP y las rutinas de destete fueron pequeñas, pero el impacto significativo de la interacción RC x destete en todos los parámetros fue de particular interés, y fue particularmente grande en el caso de la eficiencia de las emisiones cuando el LW final (FLW) del las vacas fueron consideradas. Las estimaciones de las emisiones totales de CH4 (kg) durante el período de gestación indican que, independientemente del RC, las vacas CW y EW eran emisores iguales (37.86 ± 0.506 vs 37.47 ± 0.476), mientras que los valores agrupados en el primero (37.38 ± 0.383) y el segundo ( 37.94 ± 0.390) RC también fueron similares.
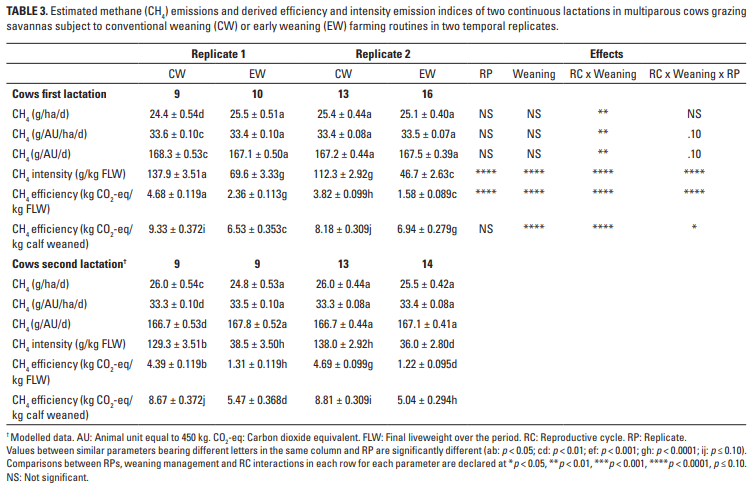
Los perfiles de emisión de CH4 de la vaca durante los dos períodos de lactancia se presentan en la Tabla 3. Los resultados muestran que el modelo pronosticó efectos consistentes y significativos de la interacción entre RC y las prácticas de destete en términos de emisiones diarias (p <0.01) o expresiones de índices (p <0,0001). Sin embargo, los índices de intensidad y eficiencia pronosticados teniendo en cuenta el FLW de las vacas sobre las lactancias se vieron significativamente afectados (p <0,0001) por todos los efectos considerados. Promediada sobre las lactaciones, hubo una diferencia grande y prácticamente importante (p <0.0001) en las emisiones totales de CH4 entre vacas CW (38.67 ± 0.456 kg) y EW (14.40 ± 0.435 kg), mientras que los valores entre las primeras (26.74 ± 0.412 kg) y la segunda lactancia (26.33 ± 0.424 kg) fueron similares.
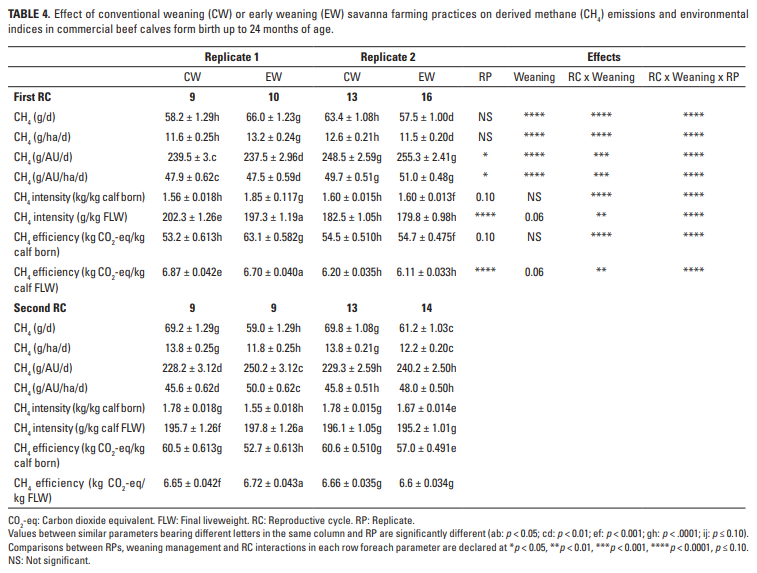
Los resultados desde el nacimiento hasta la edad de un año indican que todos los parámetros de las emisiones de CH4 se vieron afectados por la interacción de RC y los tratamientos de destete, pero nuevamente la inclusión del efecto RP en la interacción fue dominante (p <0,0001; Tabla 4). En paralelo, el impacto ambiental completo fue mayor (p <0.001) en CW (43.11 ± 0.498 kg CH4) que en EW (40.27 ± 0.472 kg CH4). De manera similar, en relación con el primer RC (39.59 ± 0.402 kg), las emisiones de CH4 fueron mayores en el segundo RC (43.79 ± 0.410 kg).
DISCUSIÓN
El análisis retrospectivo de esta investigación del flujo de CH4 derivado de LWderived fue diseñado para determinar el impacto del rendimiento extensivo del rebaño de vacas en las emisiones entéricas de CH4 y potencialmente contribuir a las oportunidades de reducción al considerar prácticas alternativas de cría de rebaños. El hallazgo más significativo fue la interacción entre los CR y las prácticas de manejo de destete de la agricultura como un factor crítico para asociar dinámicamente la producción extensiva de carne de vacuno y el conocimiento ambiental. Este resultado está en línea con Donoghue et al. (2016) que indicaron que en el ganado vacuno se pueden requerir múltiples mediciones de emisiones de CH4 en cada animal a mediano y largo plazo para registrar con precisión los rasgos de CH4 como DMI, emisiones diarias de CH4 (g) y rendimiento de CH4 (g / kg DMI) .
Confirmando los hallazgos de emisión de CH4 de Ku-Vera et al. (2018) y al reflexionar sobre la escala de nuestra representación virtual, nuestro modelo matemático ofrece huellas de carbono confiables y detalladas para promover la combinación de decisiones agrícolas tácticas. Esto sugiere qué prácticas de gestión seguir, qué clases de acciones mantener, cuándo destetar, qué vender y cuándo vender para ser más productivos de manera más eficiente y sostenible. En este sentido, como lo sugiere Tedeschi (2019), la integración de más conceptualizaciones mentales, investigación de campo y dominios virtuales utilizando cuerpos locales de datos y conocimiento científico previamente acumulados permite líneas de investigación complementarias. Por lo tanto, el conocimiento sistémico local detallado del amplio impacto ambiental de la carne de res debería respaldar una mejora potencial en la precisión del inventario de gases de efecto invernadero (IDEAM 2016) de Colombia en relación con las condiciones fisiológicas dinámicas de los animales. Para el mejor conocimiento de los autores, el inventario de ganado de GEI de Colombia se basa en un enfoque estándar de Nivel 1 del Panel Intergubernamental sobre el Cambio Climático [IPCC (2006)] que no considera los factores de emisión de CH4 medidos de ganado nativo o adaptado a los trópicos locales bajos pero se derivan de escenarios templados internacionales (56 kg CH4 / cabeza por año).
Colectivamente, nuestras observaciones están de acuerdo con el estudio de clúster y metaanálisis de Cottle y Eckard (2018) que señaló que para los informes de inventario de GEI, se requieren estimaciones específicas de cada país de las emisiones diarias y de rendimiento de CH4 a partir de los datos generados en cada país en lugar de eso derivado del método estándar de la cuenta de GEI del IPCC. Como señalaron Cottle y Eckard (2018), una serie de razones pueden contribuir a las discrepancias entre los enfoques de contabilidad mundial y local, que incluyen, entre otros, el marco de interacción fisio-metabólico adoptado en este documento.
Esas discrepancias más las diferencias significativas entre las dietas templadas y tropicales en la ingesta de energía metabolizable y la relación de ingesta de energía digestible a lo largo del año son probablemente indicativas de mediciones de CH4 más precisas en nuestro estudio, mientras que la evidencia que respalda el hecho también está representada por el cambio dinámico de LW en represas y terneros para obtener emisiones precisas de DMI y CH4 entérico. El principal impulsor fisiológico de la producción de CH4 es DMI (Jonker et al.2018), pero este efecto parece estar enmascarado si se consideran los efectos digestivos de los compuestos secundarios de las plantas (RamírezRestrepo y Barry 2005; Durmic et al.2017; Vandermeulen et al. 2018)
En el contexto de las tendencias internacionales actuales y las controversias que rodean los impactos ambientales de la producción de carne de res (Henderson et al.2017), la medicina veterinaria y las ciencias animales tienen un caso elocuente para un papel más fuerte y relevante en la producción eficiente, el rendimiento financiero, sostenibilidad, discusiones globales sobre cambio climático y dentro de la investigación de emisiones y mitigación de GEI (Ramírez-Restrepo y Charmley 2015). En concierto con esta última consideración, el CH4 se encuentra en la sangre yugular de ovejas (Ramírez-Restrepo et al. 2010) y ganado bovino (Ramírez-Restrepo et al. 2016b), pero su impacto sistémico metabólico y fisiológico aún es desconocido en los rumiantes (Tedeschi et al. al.2014; Ramírez-Restrepo y Charmley 2015; Ramírez-Restrepo et al.2016a).
Tal discusión es extremadamente pertinente ya que después de la fermentación intestinal de la fibra dietética, la flora arqueológica metanogénica luminal intestinal en humanos metanogénicos convierte el gas hidrógeno en CH4, se difunde en la circulación sistémica y el gas está vinculado a un alto índice de masa corporal (es decir, obesidad; Basseri et al. 2012). Otros datos de control de casos anidados de CH4 indican que el CH4 humano está asociado con la patogénesis de enfermedades intestinales orgánicas y funcionales, que incluyen, entre otras, diverticulosis, cáncer de colon y recto, enfermedades inflamatorias del intestino y síndrome del intestino irritable (Pimentel et al. 2006; Roccarina et al.2010; Pimentel et al.2012; Rezaie et al.2017). Por lo tanto, es posible que se requiera CH4 sanguíneo intrínseco para mantener los niveles umbral y las señales antiinflamatorias, como en la activación del sistema inmune canino (es decir, leucocitos) que puede ser modulada por la ventilación con CH4 al 2.5% (Boros et al. 2012).
Colectivamente, estos datos metabólicos cuestionan si algunas de las intervenciones plausibles para reducir las emisiones de CH4 de carne en ambientes tropicales (Ramírez-Restrepo y Charmley 2015) comprometen o no el bienestar animal. En relación con una dieta basal, Ramírez-Restrepo et al. (2016b) demostraron los efectos significativos de la suplementación con saponina de semillas de té (Camellia sinensis L.) en la bioquímica del suero sanguíneo de novillos Brahman. Este argumento es apoyado por Ramírez-Restrepo et al. (2014) que encontraron en Belmont Red Composite [Africander (African Sanga) x Brahman x Hereford-Shorthorn (3/4 B. Taurus)] dirige las complicaciones terapéuticas de los sistemas digestivo, muscular y urinario cuando el polvo de arroz rojo de Monascus purpureus fermentado contiene polvo de arroz rojo una lovastatina natural (monacolina K) se complementó como un compuesto antimetanogénico.
Por lo tanto, es particularmente evidente y necesario combinar experiencias clínicas en humanos y animales para dilucidar preguntas médicas relevantes sobre interpretaciones, similitudes, sinergias y divergencias del metabolismo de CH4 y lagunas en el conocimiento. Esto debería permitir oportunidades de financiación, investigación, enseñanza, compromiso nacional e internacional. Si se acepta esta premisa, esta conectividad conduciría a redefinir aspectos de las profesiones veterinarias y de ciencias animales del futuro y su relevancia social y global para la salud en el creciente campo del cambio climático.
Los LW de las vacas cambiaron entre valores bajos al destete y LW más altos al parir, pero el ciclo fue estable y tendió a volver a los LW de concepción inicial en el siguiente RC. Este es un fenómeno repetible observado en extensos rebaños reproductores en el norte de Australia (Fordyce et al. 2013; Fordyce et al. 2014), y junto con el conocimiento actual de la sabana sugiere posibles puntos de entrada si la intención es aumentar los niveles de producción. Sin embargo, las diferencias significativas entre los sucesivos CR reflejan la extrema sensibilidad de los rebaños basados ??en la sabana a las variaciones entre años en las condiciones ambientales.
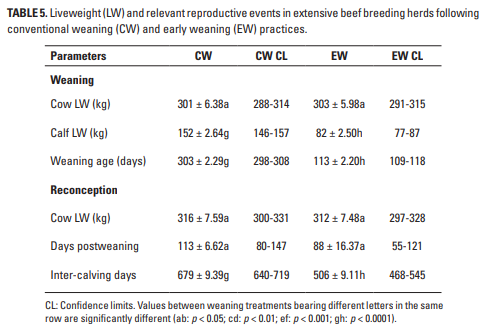
Los LW observados en la reconcepción (Tabla 5) mostraron una variabilidad moderada, según lo estimado por los límites de confianza, y no difirieron entre los tratamientos CW y EW. Estas LW son similares a las reportadas por Vera (1991; 282-300 kg) para la primera concepción de vaquillas que pastan Brachiaria humidicola, y están ligeramente por debajo de las LW en la segunda a la cuarta concepción registrada por Vera et al. (1993; 325-340 kg) para vacas que pastan ese mismo pasto. Por lo tanto, parecen ser el límite inferior para las concepciones viables llevadas a término en ganado del mismo fenotipo que se usa aquí, y extrapolando de Vera et al. (1993) también están peligrosamente cerca del LW asintótico calculado permitido en pastizales de baja calidad. Estos valores pueden compararse con los tamaños corporales maduros de vacas bien alimentadas y altamente fértiles que pastan pasturas de B. decumbens bien manejadas, que oscilaron entre 388 y 452 kg (Vera et al. 2002). La diferencia entre las vacas basadas en la sabana actualmente reportadas, y las de los animales bien alimentados en B. decumbens, y otros cultivares mejorados de Brachiaria (Peters et al. 2013), por lo tanto, indican la magnitud del desafío nutricional que enfrentan los extensos rebaños de cría. exclusivamente en sabanas nativas.
A pesar de las deficiencias minerales (Lebdosoekojo et al. 1980; Depablos et al. 2009), el bajo consumo de LW y el consumo de energía y proteínas (Ramírez-Restrepo y Barry 2005; Ramírez-Restrepo et al. 2005), y el efecto de la lactancia y la lactancia constituyen Las principales limitaciones para un rendimiento reproductivo eficiente (Osorio-Arce et al. 2002; Delgado et al. 2004; Montiel y Ahuja 2005; Schatz y Heamden 2017). Es probable que estos factores expliquen los largos períodos entre partos reportados en la Tabla 5. La gran diferencia en la duración de ese período entre los tratamientos EW y CW respalda la afirmación de que, en estas circunstancias, las largas fases de lactancia afectaron negativamente la producción de los rebaños reproductores.
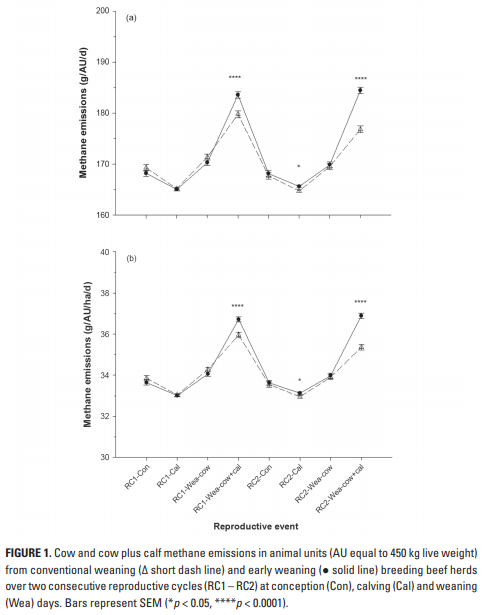
No es sorprendente que largos períodos entre partos y LW variables se asociaron con la producción total de CH4 de los RC completos, lo que lleva a bajas eficiencias ambientales por ha y por ternero (Tablas 2-4), valores que son bajos por ha solo debido a la SR baja apoyada por la sabana, pero son altas si se persigue un enfoque de sistema total como lo demuestra la representación gráfica de eventos reproductivos a lo largo del tiempo (Figuras 1a y 1b). Un contribuyente potencialmente importante a la emisión de GEI son las salidas de N de los animales de pastoreo, dado el gran efecto del N2O y otros compuestos nitrogenados (IPCC 2006), pero dados los bajos SR utilizados en las sabanas, este efecto no se tuvo en cuenta en Los resultados actuales.
CONCLUSIONES Y DIRECCIONES FUTURAS
Como la tecnología, el modelado y la variabilidad climática influyen en la forma en que las personas viven, comen, aprenden y trabajan, por su propia naturaleza, los campos de la medicina veterinaria y la ciencia animal deben adaptarse dinámicamente a las necesidades desafiantes de la producción ganadera y la presión social. Por lo tanto, en base a situaciones de granjas reales, este trabajo diferencia las estimaciones anuales de CH4 (kg / cabeza) entre RC1 y RC2 en términos de gestación más lactancia (es decir, reproducción; 47.19 ± 0.380 vs 34.75 ± 0.386; p <0.0001), destetados comerciales (14.11 ± 0.163 vs 15.25 ± 0.167; p <0.0001), stockers-año (25.12 ± 0.233 vs 26.96 ± 0.237; p <0.0001) y año (20.65 ± 0.196 vs 21.86 ± 0.199; p <0.0001). Los valores de dichos sistemas sostienen que nuestro modelo digital basado en la investigación de campo ha retratado la distancia entre las emisiones razonables de CH4 de los extensos rebaños de vacas extensas de terneros y el factor de emisión de CH4 predeterminado del IPCC latinoamericano (2006). La notable diferencia enfatiza la necesidad de mantener o avanzar cuidadosamente hacia un sistema más productivo utilizando nuestra práctica de ciencia agrícola-animal y el enfoque veterinario del ecosistema. Esto implica una comprensión sólida de la ecología del rendimiento reproductivo y la producción de carne en la interfaz de los pastizales introducidos en el país, el análisis de datos y la gestión ambiental de la granja. El enfoque combinado debería ayudar a abordar aún más la huella de carbono de los rebaños de carne en los Llanos colombianos y los determinantes sociales crecientes de la sostenibilidad y la salud en el contexto del pensamiento de los sistemas agrícolas.




{kind=link}